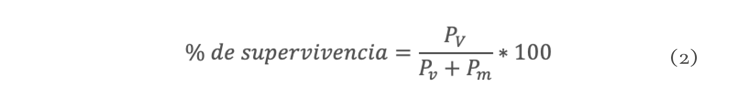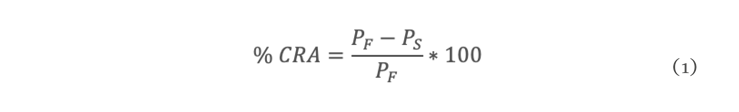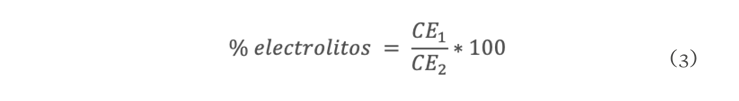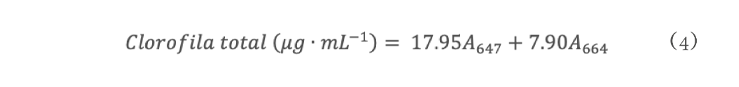Estudio del efecto de la sequía inducida por polietilenglicol en Capsicum frutescens en un sistema hidropónico
Study of the effect of drought induced by polyethylene glycol in Capsicum frutescens in a hydroponic system
María Goretty Caamal-Chan goretty_caamal@hotmail.com CONAHCyT/Centro de Investigaciones Biológicas del Noroeste S.C. La Paz, Baja California Sur, México
Rosa Isela Cruz-Rodríguez rosa.cr@tuxtla.tecnm.mx Tecnológico Nacional de México, Campus Tuxtla. Tuxtla Gutiérrez, Chiapas, México
Anayancy Lam-Gutiérrez alam@cintalapa.tecnm.mx Tecnológico Nacional de México Campus Cintalapa. Cintalapa, Chiapas, México
Nancy Ruiz-Lau nancy.rl@tuxtla.tecnm.mx CONAHCyT/ Tecnológico Nacional de México, Campus Tuxtla, Tuxtla Gutiérrez, Chiapas, México.
DOI: https://doi.org/10.31644/IMASD.34.2023.a03
Fecha de recepción: 24 de abril de 2023
Fecha de aceptación: 23 de julio de 2023
Fecha de publicación: 01 de Octubre de 2023
Agradecimientos
Al Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCyT-México) por el apoyo otorgado a través de la beca 919744 para los estudios de doctorado de Trejo-Paniagua, B. O.
Resumen
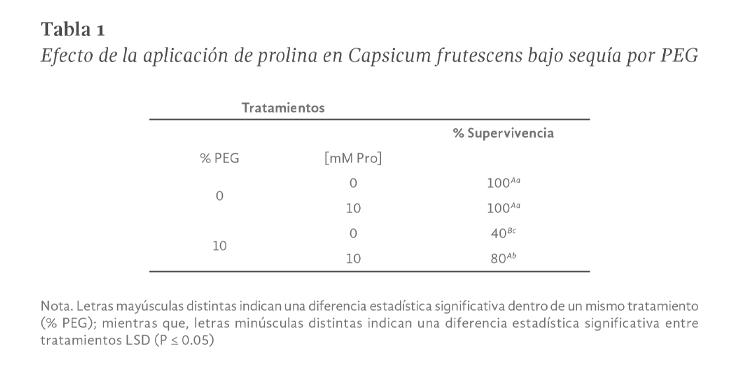
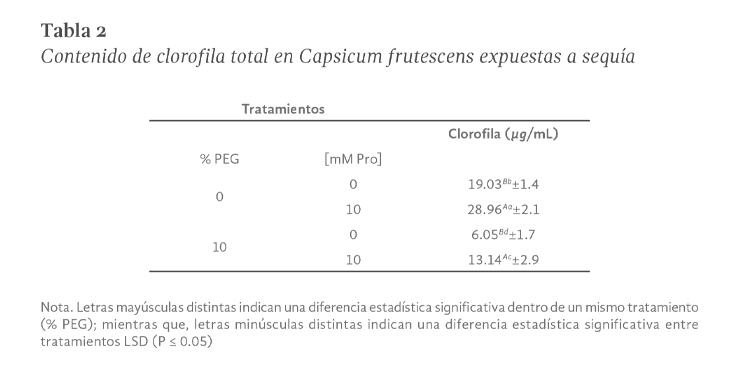
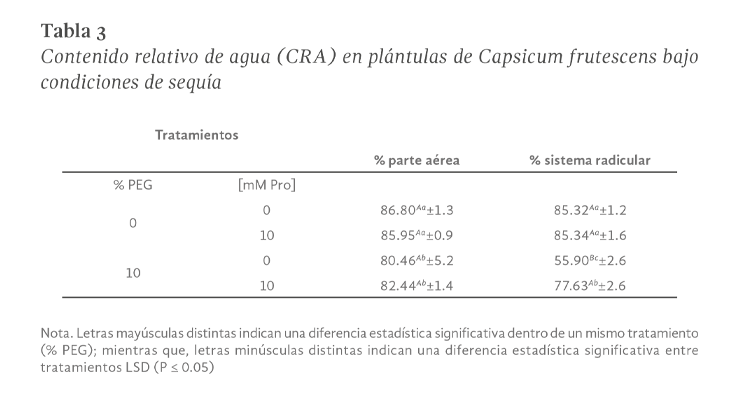
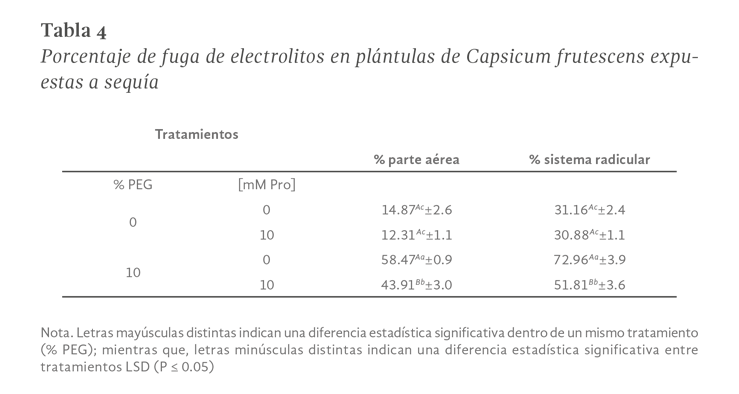
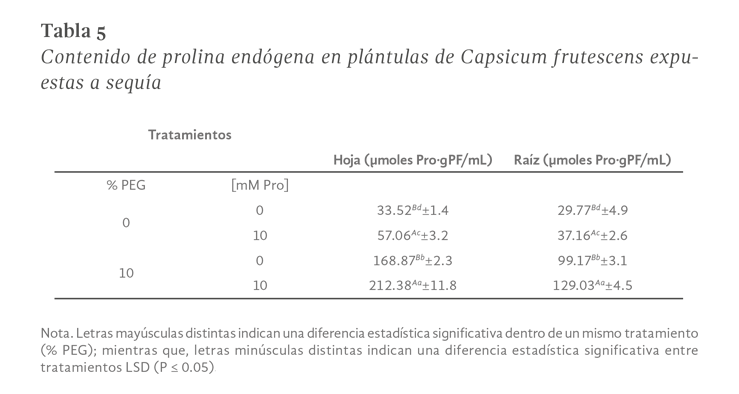
La sequía es el factor más común que limita el desarrollo y productividad de los cultivos, afectando severamente a la agricultura. En México, uno de los cultivos de importancia económica y gastronómica sensible al estrés hídrico es el género Capsicum. En el presente estudio se evaluó la aplicación de prolina en plantas de Capsicum frutescens expuestas a sequía inducida por polietilenglicol (PEG) en un sistema hidropónico. Empleando un diseño factorial 22, se evaluaron un total de 60 plántulas durante 120 h dividas en cuatro tratamientos: PEG (0 y 10 %) y Pro (0 y 10 mM) como variables de estudio. Los resultados demostraron que la exposición a 10 mM de Pro indujo un aumento significativo en la concentración de clorofila y de prolina endógena (hoja y raíz) en las plántulas en ausencia de estrés. Además, se observó un 80 % de supervivencia de las plántulas y un aumento en el contenido de prolina en aquellas que estuvieron expuestas a 10 mM Pro + 10 % PEG. De igual manera, se presentó un aumento en el contenido de clorofila (13 µg • mL-1), contenido relativo de agua (CRA) en raíz (77.6 %), porcentaje de electrolitos en hoja y raíz (~44 y ~52 % respectivamente), en comparación con el tratamiento de 0 mM Pro + 10 % PEG. Los resultados sugieren que la aplicación previa de prolina tiene un efecto positivo en la supervivencia de las plántulas bajo condiciones de sequía inducida por PEG.
Palabras clave: Sequía; Capsicum frutescens; Tolerancia; Supervivencia.
-Abstract-
Drought is the most common factor limiting crops development and productivity, severely affecting agriculture. In Mexico, one of the crops of economic and gastronomically important sensitive to water stress is the Capsicum genus. In the present study, the application of proline on Capsicum frutescens plants exposed to drought induced by polyethylene glycol (PEG) in hydroponic system was evaluated. Using a 22 factorial design, a total of 60 seedlings were evaluated for 120 hours divided into four treatments: PEG (0 and 10 %) and Pro (0 and 10 mM) as study variable. The results showed that exposure to 10 mM of Pro induced a significant increase in the chlorophyll concentration and endogenous proline (leaf and root) in seedlings in the absence of stress. In addition, 80 % survived of seedlings and an increase in proline content was observed in those exposed to 10 mM Pro + 10 % PEG. Similarly, there was an increase in chlorophyll content (13 µg • mL-1), relative water content (RWC) in the root (77.6 %), percentage of electrolytes in leaf and root (~44 and ~52 % respectively) compared to the treatment of 0 mM Pro + 10 % PEG. These results suggest that pre-application of proline has a positive effect on seedlings survival under PEG induced drought conditions
Keywords: drought; Capsicum frutescens; tolerance; survival.